












Investigadoras mendocinas del INTA EEA Mendoza, UNCuyo y Conicet estudiaron la diversidad y distribución de las cepas de B. bruxellensis en diferentes vinos -desde la uva hasta seis meses de envejecimiento- con el fin de obtener información sobre el origen y el momento de la contaminación. Los resultados demuestran que el equipamiento de la bodega puede actuar como vector de introducción de Brettanomyces, en 2 etapas críticas: el uso compartido de equipos durante la fermentación y la reutilización de barricas.

Autoras: María Elena Sturm y Selva Valeria Chimeno (INTA EEA Mendoza); Laura Analía Mercado (INTA EEA Mendoza y Facultad Ciencias Agrarias UNCuyo); Mariana Combina (INTA EEA Mendoza y Conicet); Magalí Lucía González (Facultad Ciencias Agrarias UNCuyo); María Cecilia Lerena, María Cecilia Rojo y Lucía Maribel Becerra (Conicet).
Brettanomyces bruxellensis es reconocida como la principal levadura de deterioro en los vinos tintos, produciendo fenoles volátiles que afectan negativamente la calidad del vino. Sin embargo, pocos estudios han investigado la diversidad de cepas dentro de las bodegas. Comprender la diversidad y distribución de las cepas de B. bruxellensis en diferentes vinos puede aportar información sobre el origen y el momento de la contaminación.
Este estudio tuvo como objetivo evaluar la presencia y diversidad de biotipos de B. bruxellensis durante la producción de cuatro vinos tintos en la misma bodega e identificar etapas críticas de contaminación. El análisis abarcó todo el proceso, desde la uva hasta seis meses de envejecimiento. Las levaduras de B. bruxellensis fueron aisladas e identificadas, y las cepas representativas se tipificaron mediante análisis RAPD.
Los resultados sugieren que la contaminación por B. bruxellensis no se originó a partir de una única fuente. Las uvas presentaban niveles bajos de B. bruxellensis, sin embargo, todos los vinos resultaron positivos antes del llenado en barrica. Este estudio demuestra que el equipamiento de la bodega puede actuar como vector de introducción de Brettanomyces.
Introducción
Las Brettanomyces spp. han sido descritas como la principal levadura de deterioro en el vino. Dentro de este género, la especie B. bruxellensis es un microorganismo de deterioro bien conocido en la elaboración de vino. Sin embargo, B. bruxellensis no es exclusiva del vino; también se encuentra en diversas otras bebidas alcohólicas donde no se considera una levadura de deterioro, como ciertas cervezas especiales y sidras. B. bruxellensis altera negativamente la composición química del vino al producir compuestos volátiles metabólicos que perjudican las propiedades organolépticas del producto final.
Este defecto ocurre principalmente cuando los precursores de ácidos hidroxicinámicos (ácidos p-cumárico, ferúlico y cafeico) son descarboxilados enzimáticamente por la descarboxilasa de cinamato, dando lugar a la formación de derivados vinílicos, que posteriormente son reducidos por la vinil fenol reductasa para producir derivados etílicos como 4-etilfenol (4-EP), 4-etilguayacol (4-EG) y 4-etilcatecol (4-EC). Además, otros subproductos metabólicos menores de B. bruxellensis, como los ácidos isovalérico e isobutírico, contribuyen a la percepción del “carácter Brett”. Los olores fenólicos indeseables asociados a la contaminación por Brettanomyces se han descrito con descriptores sensoriales como “medicinal”, “fenólico”, “rancio”, “sudoroso”, “ahumado”, “curita (Band-Aid®)”, “corral” o “sudor de caballo”.
Ambos nombres de esta levadura, Brettanomyces y Dekkera, se utilizan a menudo como sinónimos en la literatura científica. Además, según el Código Internacional de Nomenclatura revisado para algas, hongos y plantas (Código de Melbourne), las especies fúngicas deben asignarse a un único nombre válido. Sin embargo, la denominación Brettanomyces es más común en las industrias alimentaria y biotecnológica, donde la especie B. bruxellensis se asocia inmediatamente con el vino por todos los actores del sector. Por lo tanto, en este estudio se utiliza preferentemente el término Brettanomyces.
A diferencia de Saccharomyces cerevisiae, B. bruxellensis presenta un crecimiento lento y no es un fuerte competidor en sustratos con alto contenido de azúcar. Esta levadura tiene bajos requerimientos nutricionales y está bien adaptada a ambientes hostiles como el vino. De hecho, puede crecer en medios pobres en nutrientes, bajo condiciones de pH bajo (3–4), en presencia de altas concentraciones de etanol y con niveles elevados de sulfitos. Un ejemplo notable de esta adaptación es su papel como levadura de deterioro en el vino, donde generalmente se detecta durante el proceso de envejecimiento, aunque también puede aparecer con menor frecuencia en etapas más tempranas de la vinificación, incluyendo las uvas y el mosto.
No obstante, B. bruxellensis puede desarrollarse al final de la fermentación alcohólica, cuando otras poblaciones microbianas disminuyen, y se ha asociado principalmente con fermentaciones lentas y fermentaciones malolácticas retardadas. Aunque esta levadura no es dominante en el mosto durante la fermentación, su presencia puede afectar significativamente el producto final.
El origen de la contaminación por Brettanomyces sigue siendo incierto. Algunos autores sugieren que las uvas podrían ser una fuente de contaminación en los vinos. De hecho, los enólogos en bodegas comerciales suelen asociar viñedos específicos con la presencia recurrente de Brettanomyces en los vinos elaborados a partir de sus uvas. Sin embargo, debido a las dificultades para aislar B. bruxellensis a partir de uvas, esta hipótesis sigue siendo difícil de confirmar. Por otro lado, varios estudios proponen que la propia bodega es la principal fuente de contaminación, siendo la reutilización de barricas de roble el punto de contaminación más frecuentemente identificado. Además, se han descrito otras superficies contaminadas con B. bruxellensis en la bodega, como las paredes de la cava, desagües, bombas, equipos de trituración y cualquier tanque o línea de trasiego que sea difícil de sanitizar eficazmente.
La contaminación de los vinos por B. bruxellensis ha aumentado en los últimos años debido a la evolución de las técnicas de vinificación que favorecen la producción de vinos con maceraciones prolongadas, lo que incrementa la extracción de precursores, así como vinos que pueden no estar sulfitados, no filtrados, criados sobre lías o madurados durante períodos más largos en barrica. Todos estos factores se consideran favorables para el crecimiento de B. bruxellensis.
Las estrategias para controlar el crecimiento de esta levadura de deterioro representan un desafío significativo para los enólogos. Comprender la diversidad de cepas y su distribución en diferentes vinos puede ayudar a revelar el origen y el momento de la contaminación, permitiendo diseñar estrategias de prevención y control.
Materiales y Métodos
Viñedos y condiciones de vinificación
Se seleccionaron cuatro viñedos ubicados en la provincia de Mendoza (Argentina) para este estudio. Los viñedos deMerlot (Me) y Cabernet Sauvignon (Cs) estaban situados en Agrelo, Luján de Cuyo, dentro de la región vitivinícola conocida como “Zona Alta del Río Mendoza”, mientras que los dos viñedos de Malbec (Mb2 y MbC) se encontraban en Villa Bastías, Tupungato, dentro de la región vitivinícola denominada “Valle de Uco”. Ambas regiones pertenecen al grupo climático IH + 2 IF + 1 IS + 1, caracterizado por un clima cálido con noches frías y sequía moderada. El clima es seco, con precipitaciones anuales cercanas a 200 mm.
Los viñedos pertenecían a una bodega comercial donde se llevó a cabo el proceso de vinificación. La bodega estaba ubicada a una distancia considerable de los viñedos muestreados, lo que evitó cualquier posible contaminación antropogénica cruzada entre viñedos y bodega. Las muestras de uva se tomaron de viñedos destinados a producir vinos de calidad media a alta. Las vides estaban conducidas en un sistema de espaldera baja con adecuado control de enfermedades, lo que dio lugar a uvas de alta calidad sanitaria.
Las uvas fueron trituradas y se obtuvo mosto fresco. Se realizó un sulfitado inicial utilizando metabisulfito de potasio para alcanzar una dosis total final de SO₂ de 30 ppm en el mosto. La fermentación tuvo lugar en tanques de acero inoxidable de 250 hL, inoculados con una cepa comercial de Saccharomyces cerevisiae (Lalvin EC1118, Lallemand Inc., Montreal, Canadá). La fermentación maloláctica se llevó a cabo de forma espontánea. Ambas fermentaciones se realizaron a 26 ± 2 °C.
Posteriormente, el vino se transfirió a barricas de roble de 200 L para su envejecimiento durante seis meses. Durante este período, las barricas se mantuvieron en una sala acondicionada a 15 °C y 75–80% de humedad. Para mantener los niveles de SO₂ durante la crianza, la bodega emplea dos prácticas: la adición individual de SO₂ a cada barrica, o la práctica conocida como “trasiego y SO₂”, donde el vino se extrae de las barricas y se coloca en un tanque, se corrige el nivel de SO₂ y luego se devuelve a las barricas. Esta práctica combina vinos provenientes de barricas usadas por un lado y de barricas nuevas por otro.
Muestreo de uvas y vinos
En cada viñedo se muestrearon diez plantas a lo largo de dos transectas diagonales, recolectando tres racimos sanos por planta. En total, se consideraron 30 racimos por viñedo. Para aumentar la probabilidad de detectar Brettanomyces, también se recolectaron muestras adicionales de uvas dañadas (deshidratadas, podridas, decoloradas o manchadas), así como uvas afectadas por Planococcus ficus y racimos ubicados cerca de los bordes del viñedo, próximos a árboles. Estas muestras se analizaron por separado.
Todas las muestras se colocaron en bolsas plásticas y se almacenaron a 4 °C hasta su análisis en laboratorio. Los mostos de los cuatro viñedos se fermentaron en la bodega comercial y se monitorearon para detectar la presencia de Brettanomyces desde el estrujado hasta el final del envejecimiento en barrica. Se tomaron muestras de 250 mL en recipientes estériles a lo largo de todo el proceso.
Durante el envejecimiento, se seleccionaron aleatoriamente cinco barricas, que fueron marcadas y monitoreadas durante seis meses. Además, se incluyeron cinco muestras de otros vinos positivos a Brettanomyces en crianza en la misma bodega. Las muestras se identificaron como: inicio de fermentación (BF), mitad de fermentación (MF), fin de fermentación o 0° Baumé (0Be), vino antes de barrica (BB), envejecimiento en barrica mes 1 (M1) hasta mes 6 (M6), y vinos positivos a Brettanomyces en barrica: 16088, 16091, 7042, 7090 y 7129.
Detección y aislamiento de Brettanomyces
Las bolsas refrigeradas con uvas se abrieron en el laboratorio bajo condiciones estériles y se trituraron asépticamente. Se colocaron 45 mL de uvas trituradas en un matraz estéril de 100 mL y se añadió 1 mL de una solución de enriquecimiento selectiva y diferencial para Brettanomyces (BSDES) 50X. Esta solución contenía: extracto de levadura (100 g/L), KNO₃ (15 g/L), ácido p-cumárico (5 g/L), cloranfenicol (25 g/L) y cicloheximida (10 g/L). Además, se agregaron 4 mL de etanol (99%) para aumentar la selectividad del medio.
Los matraces se incubaron a 28 °C con agitación suave (150 rpm) durante 60 días, realizando muestreos cada 10 días. Tras el enriquecimiento, las células viables se sembraron en placas con medio selectivo diferencial (BSDM), compuesto por glucosa, extracto de levadura, agar, ácido p-cumárico, cloranfenicol y cicloheximida, con pH ajustado a 4,7. Después de la esterilización, se añadió 6% de etanol.
Las colonias se consideraron positivas para Brettanomyces si mostraban crecimiento después de cinco días, producción de olor fenólico y morfología celular típica bajo microscopía. En algunos casos también se evaluó la producción de ácido en medio WLN. Las colonias representativas se aislaron, purificaron y conservaron en glicerol al 30% a −80 °C para su posterior identificación molecular.
Identificación de especies mediante RFLP-PCR
La extracción de ADN se realizó a partir de cultivos puros activos. Las células se cultivaron en medio líquido YPD y se incubaron durante 48 h a 28 °C. La extracción de ADN siguió un método modificado de Hoffman y Winston, incluyendo lisis celular, purificación con fenol/cloroformo/alcohol isoamílico y precipitación con etanol.
La integridad del ADN se evaluó mediante electroforesis en gel de agarosa al 0,7%, y la cuantificación se realizó con espectrofotómetro NanoDrop. La identificación molecular a nivel de especie se llevó a cabo mediante RFLP-PCR utilizando cebadores específicos del género Dekkera, seguido de digestión con la enzima de restricción DdeI para diferenciar D. bruxellensis de D. anomala.
Tipificación molecular de aislados de B. bruxellensis
Los aislados se analizaron a nivel de cepa mediante RAPD utilizando tres cebadores: M13, COC y OPK03. Para evaluar el poder discriminatorio de los marcadores, se tipificaron inicialmente siete cepas de referencia previamente identificadas como B. bruxellensis, incluyendo seis cepas locales de la colección CoMIM (INTA Mendoza) y una cepa de referencia de la colección española (CECT11045).
Posteriormente, estos marcadores se utilizaron para genotipificar 85 aislados representativos previamente identificados como B. bruxellensis.
Análisis de conglomerados (cluster)
Los patrones de bandas obtenidos tras la electroforesis se normalizaron utilizando el software PyElph versión 1.4 para construir una matriz de presencia/ausencia. Se calculó el coeficiente de Dice para estimar la similitud entre los aislados.
Se construyó un dendrograma utilizando el método UPGMA con los programas PAST 3.21 y FIG TREE versión 1.4.4. El coeficiente de correlación cofenética se calculó como medida de la fiabilidad del agrupamiento utilizando el mismo software.
Resultados
Presencia de Brettanomyces en uvas
La dificultad para aislar Brettanomyces a partir de uvas es ampliamente reconocida. Por esta razón, se realizó un muestreo extensivo de uvas. Se tomaron muestras representativas de los viñedos y se recolectaron distintos tipos de uvas. Las muestras se clasificaron como uvas sanas, uvas dañadas, uvas deshidratadas, uvas decoloradas o manchadas, uvas recolectadas en zonas sombreadas cercanas a árboles y uvas dañadas por Planococcus ficus.
Los resultados mostraron que solo una muestra de uvas presentó presencia de Brettanomyces, a pesar de haberse realizado 24 cultivos de enriquecimiento durante un período de 60 días. Las uvas dañadas del viñedo Malbec C (MbC) mostraron un recuento de 1,5 × 10⁴ UFC/mL después del enriquecimiento.
Presencia de Brettanomyces durante la vinificación
Los mostos de los cuatro viñedos estudiados fueron fermentados en una bodega comercial utilizando tanques de acero inoxidable individuales. La presencia de Brettanomyces se monitoreó desde el estrujado hasta el final del envejecimiento.
Para seguir las poblaciones de Brettanomyces en cada uno de los vinos producidos y comprender el impacto de las prácticas de vinificación empleadas, los resultados se presentan por separado para cada vino.
Merlot (Me)
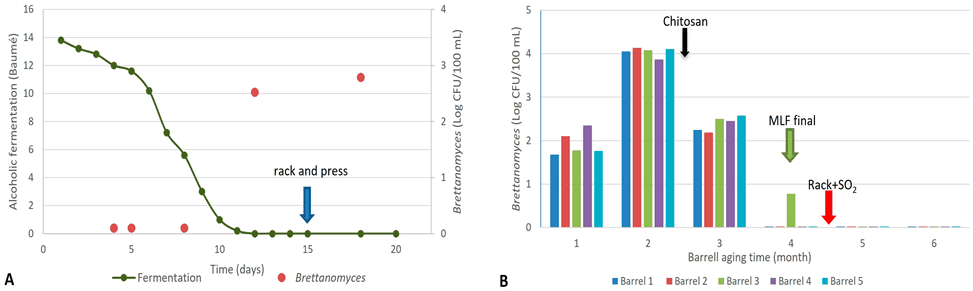
En la fermentación realizada con uvas del viñedo de cv. Merlot (Me), no se encontraron muestras positivas para Brettanomyces durante las primeras etapas de la fermentación alcohólica. Sin embargo, la muestra tomada al final de la fermentación alcohólica mostró resultados positivos, con poblaciones de 3,3 × 10² UFC/100 mL. Estas poblaciones continuaron aumentando, alcanzando 6,2 × 10² UFC/100 mL en la muestra de vino recolectada pocos días antes del llenado en barrica (Figura 1A).
Posteriormente, el vino se colocó en barricas sin adición de SO₂, ya que la fermentación maloláctica espontánea aún no había finalizado. La Figura 1B muestra los recuentos de Brettanomyces en las cinco barricas seleccionadas y monitoreadas durante seis meses de envejecimiento. Se observó que las barricas presentaron altos recuentos de Brettanomyces durante los primeros tres meses.
En el primer mes, las poblaciones fueron similares a las observadas antes del llenado en barrica. Sin embargo, en el segundo mes, las poblaciones aumentaron aproximadamente dos órdenes logarítmicos, alcanzando valores cercanos a 10⁴ UFC/100 mL. Tras este incremento, la bodega aplicó quitosano (No Brett Inside®, Lallemand Inc.) a una dosis de 4 g/hL, lo que redujo las poblaciones de Brettanomyces en 1,66 unidades logarítmicas en las muestras del tercer mes de envejecimiento.
Para el cuarto mes, las poblaciones eran indetectables o muy bajas (Figura 1B). Cabe destacar que, antes de la toma de muestras en ese momento, la bodega añadió SO₂ (33 ppm), lo que pudo haber potenciado el efecto del quitosano, manteniendo las poblaciones cultivables de Brettanomyces en niveles indetectables (<1 UFC/100 mL) hasta el final del período de envejecimiento.
Cabernet Sauvignon (Cs)
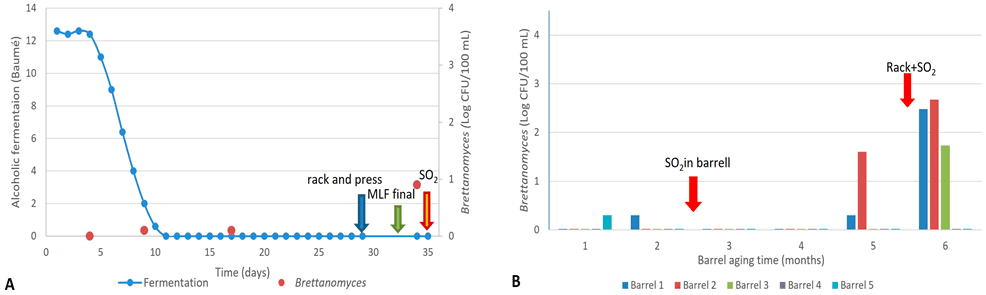
No se observaron recuentos positivos de Brettanomyces durante la fermentación alcohólica ni maloláctica en el Cabernet Sauvignon producido en la bodega. Poco después de завершar la fermentación maloláctica, pero antes de la adición de SO₂ (30 ppm) previa al llenado en barrica, se observó un resultado positivo para Brettanomyces, aunque con un recuento bajo (8 UFC/100 mL) (Figura 2A).
Durante el envejecimiento en barrica, los vinos mostraron recuentos negativos o muy bajos de Brettanomyces (2 UFC/100 mL) hasta el quinto mes. Al final del segundo mes de crianza, la bodega ajustó los niveles de SO₂ (36 ppm) mediante la adición directa a cada barrica.
En el quinto mes de envejecimiento, dos barricas mostraron presencia de Brettanomyces, con poblaciones que aumentaron en el mes siguiente, junto con la aparición de nuevas barricas contaminadas, superando valores de 10² UFC/100 mL en dos de ellas (Figura 2B).
Es importante destacar que entre el quinto y el sexto mes de crianza, la bodega realizó la práctica conocida como “trasiego y SO₂”, tal como se describió previamente en la sección de materiales y métodos. Los resultados sugieren que esta práctica puede favorecer la diseminación de la contaminación por Brettanomyces.
Malbec 2 (Mb2)
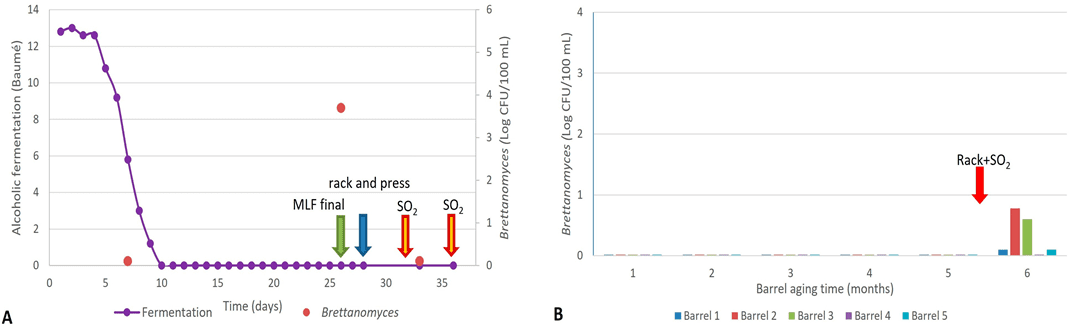
Durante la fermentación alcohólica del mosto de Malbec 2 (Mb2), no se detectó la presencia de Brettanomyces. Sin embargo, una vez finalizada la fermentación, el vino se mantuvo en el tanque hasta que concluyó la fermentación maloláctica espontánea (FML) en un período de 19 días.
Una muestra de vino tomada al final de la FML mostró recuentos elevados de Brettanomyces, con poblaciones de 3,3 × 10³ UFC/100 mL. Posteriormente, el vino fue trasegado y prensado, y se añadió SO₂ (30 ppm). Estas acciones redujeron las poblaciones de Brettanomyces en 1,5 unidades logarítmicas, pero aún se mantuvieron poblaciones viables de 1 × 10² UFC/100 mL.
Como medida correctiva, la bodega volvió a añadir SO₂ (25 ppm) antes del llenado en barrica (Figura 3A). No se observaron muestras positivas de Brettanomyces durante los primeros cinco meses de envejecimiento en barrica. Sin embargo, a partir del sexto mes, la mayoría de las barricas mostraron recuentos positivos de esta levadura, con poblaciones superiores a 10² UFC/100 mL (Figura 3B).
Cabe destacar que entre el quinto y el sexto mes de crianza, la bodega realizó en este vino la práctica de “trasiego y SO₂”, mencionada anteriormente. Estos resultados resaltan nuevamente el impacto de esta práctica en la diseminación de la contaminación por Brettanomyces.
La única barrica (B5) que no presentó recuentos positivos durante el sexto mes correspondió a un vino al que se le había añadido quitosano antes de volver a llenar la barrica (Figura 3B).
Malbec C (MbC)
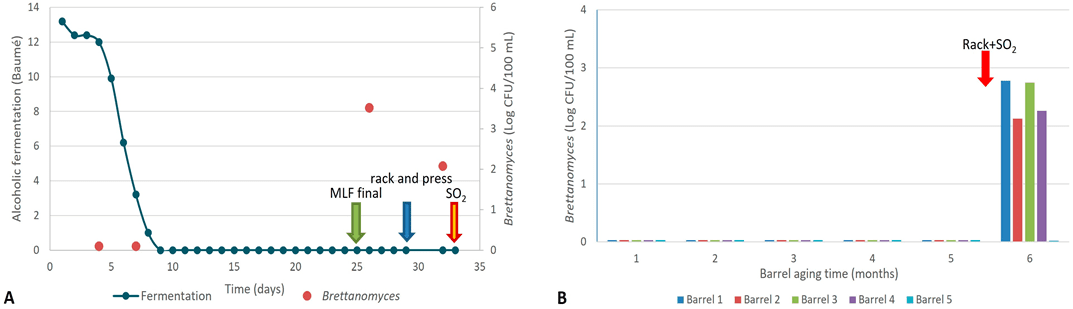
Las muestras provenientes de la fermentación de los mostos de Malbec C no mostraron presencia de Brettanomyces. Esto resultó contrario a lo esperado, ya que cabe destacar que las uvas de este viñedo fueron las únicas que dieron un resultado positivo para el aislamiento de Brettanomyces.
Por otro lado, la muestra tomada al final de la fermentación maloláctica espontánea (16 días después de finalizada la fermentación alcohólica) presentó poblaciones de aproximadamente 5 × 10³ UFC/100 mL en el vino, las cuales se redujeron significativamente tras el sulfitado realizado durante el trasiego y prensado (Figura 4A).
El seguimiento de las poblaciones de Brettanomyces durante el envejecimiento de este vino mostró recuentos negativos durante los primeros cinco meses de crianza. De manera similar a los resultados obtenidos para Malbec 2 (Mb2), los vinos presentaron recuentos positivos de Brettanomyces en el sexto mes de envejecimiento, tras la aplicación de la práctica de “trasiego y SO₂” por parte de la bodega (Figura 4B).
Un análisis general de los cuatro vinos estudiados reveló que todos presentaron poblaciones de Brettanomyces antes del llenado en barrica. Los vinos mostraron recuentos positivos principalmente durante la fermentación maloláctica espontánea. Antes del envejecimiento en barrica, la bodega aplica sulfitado, lo que ayuda a mantener bajas las poblaciones cultivables de Brettanomyces. Sin embargo, la posterior práctica de “trasiego y SO₂” promueve un aumento y la diseminación de las poblaciones de Brettanomyces dentro de las barricas.
Identificación molecular y tipificación de aislados de Brettanomyces
Para comprender la diversidad de las poblaciones de Brettanomyces en las muestras positivas recolectadas y, en consecuencia, identificar el origen y el punto de contaminación durante la vinificación y el envejecimiento, los aislados se identificaron primero a nivel de especie y luego se tipificaron molecularmente.
El análisis de restricción de los amplicones obtenidos mediante PCR con el par de cebadores específicos DB90F/DB394R, utilizando la enzima de restricción DdeI, mostró dos fragmentos de aproximadamente 154 pb y 129 pb, confirmando así que todos los aislados pertenecían a B. bruxellensis.
La tipificación molecular de los aislados de B. bruxellensis se realizó utilizando los marcadores RAPD M13, COC y OPK03. En primer lugar, para evaluar el poder discriminatorio de los marcadores seleccionados, se amplificaron siete cepas de referencia. Los marcadores utilizados generaron perfiles moleculares con entre 8 y 15 bandas de diferentes pesos moleculares (Figura 5A–C).
Los dendrogramas UPGMA mostraron distintos grados de relación entre las cepas, dependiendo del marcador molecular utilizado (Figura 5D–F). Los marcadores M13 y COC RAPD produjeron agrupamientos similares para la mayoría de las cepas, con la cepa de referencia española mostrando una relación más distante. En cambio, el marcador OPK03 RAPD generó un agrupamiento completamente diferente, ubicando la cepa de referencia española dentro de un clúster junto con cepas locales.
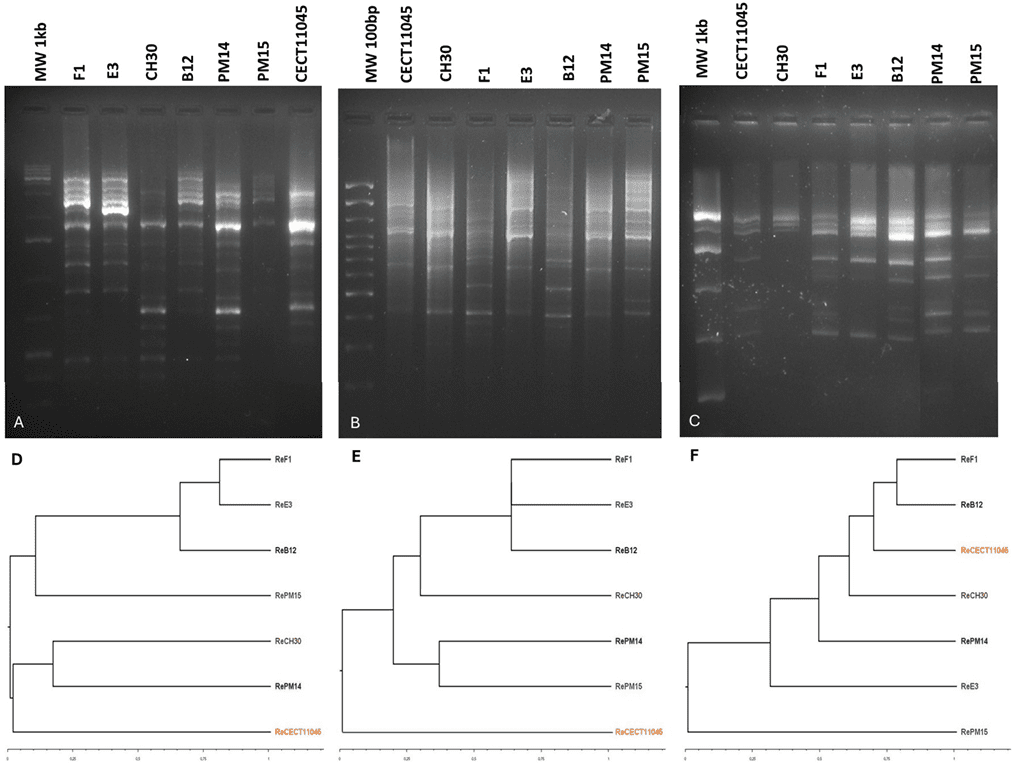
A pesar de esta discrepancia, el marcador OPK03 RAPD se incluyó en la tipificación de los aislados en este estudio, ya que demostró un buen poder discriminatorio y había sido recomendado para este tipo de análisis por Oro et al.
Posteriormente, estos tres marcadores moleculares se utilizaron para tipificar 72 aislados de B. bruxellensis provenientes de los cuatro viñedos analizados —incluyendo aislados desde las uvas hasta seis meses de envejecimiento del vino—, así como 13 aislados de cinco vinos contaminados de la misma bodega.
Se construyó una matriz binaria combinada incorporando las bandas generadas por los tres marcadores, dando lugar a una matriz de presencia/ausencia con un total de 178 bandas (54 para M13, 59 para COC y 65 para OPK03). El nivel de discriminación entre los aislados fue alto, con la primera ramificación del dendrograma ocurriendo a un coeficiente de similitud inferior a 0,05 (Figura 6).
La tipificación global mediante la combinación de los marcadores moleculares reveló la existencia de 20 clústeres genéticos a un coeficiente de similitud de 0,30 (Figura 6).
En el análisis del dendrograma UPGMA (Figura 6), se puede observar inicialmente la presencia de varios clústeres que agrupan muestras de una misma variedad de uva en la misma etapa de envejecimiento. El ejemplo más claro se observa en cuatro clústeres que agrupan aislados obtenidos durante el sexto mes de crianza.
El clúster 1 agrupó cinco aislados del vino Cabernet Sauvignon (CsM6), el clúster 6 agrupó 11 aislados de vinos Malbec C y Malbec 2 (MbCM6 y Mb2M6), y los clústeres 7 y 8 agruparon seis y dos aislados del vino Malbec 2 (Mb2M6), respectivamente.
Un análisis más detallado de estas muestras reveló además que los aislados dentro de cada clúster provenían del mismo vino envejecido en diferentes barricas. Asimismo, los vinos durante el envejecimiento en barrica mostraron la presencia de múltiples biotipos de B. bruxellensis.
Por ejemplo, los aislados de Merlot se distribuyeron en seis clústeres diferentes (clúster 3, 4, 10, 15, 16 y 19); los de Cabernet Sauvignon en cinco clústeres (clúster 1, 3, 5, 14 y 15); los de Malbec C en tres clústeres (clúster 6, 11 y 20); y los de Malbec 2 en cuatro clústeres (clúster 6, 7, 8 y 17) (Figura 6). Este hallazgo confirma que las poblaciones de B. bruxellensis en los vinos durante el envejecimiento están compuestas por múltiples cepas.
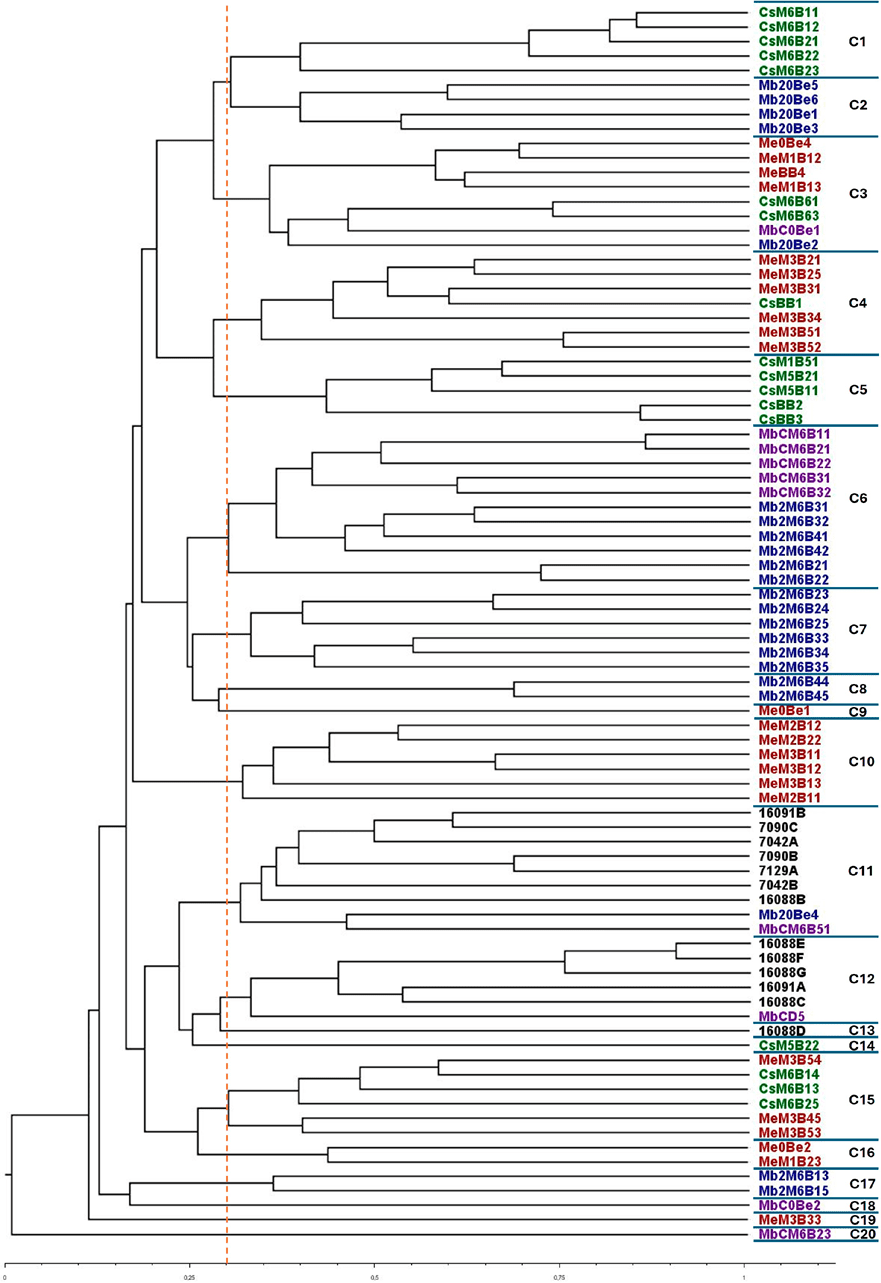
Por otro lado, otro clúster agrupó aislamientos provenientes de vinos durante la fermentación y el primer mes de envejecimiento. Esta agrupación de aislamientos muestra que las cepas presentes durante la fermentación pueden persistir en el vino una vez que es transferido a la barrica. Por ejemplo, el clúster 16 agrupó dos aislamientos del final de la fermentación (Me0Be) y del primer mes de envejecimiento (MeM1) para el vino Merlot.
Además, el clúster 3 agrupó ocho aislamientos de vinos de diferentes varietales durante la fermentación, incluyendo cuatro aislamientos de Merlot al final de la fermentación (Me0Be), antes del llenado de barricas (MeBB) y durante el primer mes de envejecimiento (MeM1), así como dos aislamientos de Malbec C y Malbec 2 al final de la fermentación (MbC0Be y Mb20Be). La presencia de aislamientos provenientes de muestras de fermentación de diferentes vinos (Merlot, Malbec C y Malbec 2) sugiere cierto nivel de diseminación de cepas entre los tanques de fermentación, potencialmente mediada por equipos compartidos en la bodega.
Un análisis más profundo de los aislamientos del vino Merlot, que mostró contaminación con B. bruxellensis desde la fermentación hasta el tercer mes de envejecimiento, revela que, como se mencionó previamente, los aislamientos de la fermentación reaparecieron durante el primer mes de envejecimiento (Figura 6, clústeres 3 y 16). Sin embargo, los aislamientos de este vino durante el segundo y tercer mes de envejecimiento se agruparon en diferentes clústeres.
El clúster 4 agrupó seis aislamientos de Merlot en el tercer mes de envejecimiento (MeM3), y el clúster 10 agrupó seis aislamientos del mismo vino correspondientes al segundo y tercer mes de envejecimiento (MeM2 y MeM3). Este hallazgo sugiere que las diferentes cepas de B. bruxellensis detectadas durante el envejecimiento pueden haberse originado a partir de barricas reutilizadas o de poblaciones preexistentes que permanecieron indetectables debido a su bajo número inicial.
Este mismo comportamiento también puede observarse al analizar los aislamientos de Cabernet Sauvignon. Aunque este vino mostró un bajo nivel de contaminación, con solo unas pocas barricas contaminadas durante el envejecimiento, los aislamientos del vino antes del llenado de barricas (CsBB) y durante el envejecimiento (CsM1 y CsM5) se agruparon juntos en el clúster 5. Sin embargo, después de la práctica de “trasiego y SO₂” realizada por la bodega antes del sexto mes de envejecimiento, los aislamientos de B. bruxellensis se agruparon en diferentes grupos, como el clúster 1 (5 aislamientos), el clúster 3 (2 aislamientos) y el clúster 15 (3 aislamientos) (Figura 6). Este hallazgo confirma una vez más que las cepas presentes en vinos contaminados podrían permanecer en las barricas durante el envejecimiento, y que la práctica de “trasiego y SO₂” podría dispersar la contaminación e introducir nuevas cepas.
Los 13 aislamientos de B. bruxellensis obtenidos de los cinco vinos contaminados durante el envejecimiento proporcionados por la bodega (16088; 16091; 7042; 7090; y 7129) se agruparon en dos clústeres relacionados (clúster 11 y clúster 12) y se separaron de los aislamientos de los cuatro vinos estudiados (Me, Cs, MbC y Mb2). Curiosamente, el único aislamiento de uvas de Malbec dañadas (MbCD5) fue incluido en el clúster 12 (Figura 6).
Discusión
La detección de Brettanomyces en el presente estudio se llevó a cabo utilizando medios de cultivo; por lo tanto, los resultados reflejan poblaciones cultivables, que en consecuencia son metabólicamente activas. Es ampliamente conocido que un estado viable pero no cultivable (VBNC) en Brettanomyces puede ser inducido por la adición de SO₂. Sin embargo, estudios previos no han establecido claramente si la producción de fenoles volátiles está vinculada al estado fisiológico, a la fase de crecimiento de B. bruxellensis, o a ambos, y los hallazgos han sido a menudo contradictorios.
Algunos estudios sugieren que la actividad enzimática en células VBNC de Brettanomyces podría dar lugar a defectos sensoriales en el vino. No obstante, estos estudios se han realizado principalmente sobre poblaciones altamente activas antes de la inducción del estado VBNC.
Nuestro estudio previo demostró claramente la relación entre la capacidad de Brettanomyces para crecer en el vino y alcanzar poblaciones críticas, lo que conduce a la aparición de defectos sensoriales en los vinos. Además, Godoy et al. demostraron que ambas enzimas responsables de la producción de fenoles presentan cierta inestabilidad en presencia de etanol, con una marcada disminución de la actividad de la p-cumarato descarboxilasa (CD).
Es importante destacar que, en éste estudio, los recuentos de Brettanomyces cultivables en los vinos analizados no superaron el umbral crítico de población (1000 células/mL), el cual se asocia con la aparición de defectos sensoriales perceptibles en un corto período. En consecuencia, incluso si hubiera existido actividad enzimática residual en las células VBNC en nuestro estudio, la baja población de Brettanomyces detectada antes de la adición de SO₂ fue insuficiente para producir fenoles volátiles por encima del umbral de percepción. Como resultado, los vinos de este estudio no desarrollaron defectos organolépticos.
Nuestros hallazgos confirmaron que B. bruxellensis fue la única especie perteneciente al género Brettanomyces asociada con el vino y la bodega; este hecho ya ha sido demostrado previamente. Diversos estudios han explorado la diversidad genética de B. bruxellensis utilizando diferentes técnicas de “fingerprinting” como RAPD, polimorfismo de longitud de fragmentos amplificados (AFLP), marcadores microsatélites, análisis de restricción de ADN mitocondrial (mtDNA) y otras. Estos estudios destacan una importante diversidad intraespecífica de B. bruxellensis, lo que dificulta la predicción de su aparición y comportamiento en fermentaciones industriales.
En el estudio, la construcción de una matriz binaria combinando los tres marcadores utilizados para la tipificación molecular dio como resultado un conjunto de datos con un alto número de bandas. La elevada distancia molecular observada entre los aislamientos en este estudio es consistente con investigaciones previas que han demostrado que la técnica RAPD puede generar variabilidad entre perfiles moleculares de cepas, incluso cuando las concentraciones de ADN y los reactivos están estandarizados. Para la interpretación de los resultados, los aislamientos agrupados dentro del mismo clúster fueron considerados biotipos genéticamente cercanos, lo que sugiere que estos aislamientos podrían ser variantes de la misma cepa o de un grupo de cepas estrechamente relacionadas.
A pesar de los esfuerzos por aislar Brettanomyces a partir de uvas, solo una muestra resultó positiva tras un período de enriquecimiento de 60 días. Se obtuvo un único aislamiento a partir de uvas dañadas sin signos visibles de podredumbre. Este hallazgo limitó nuestra capacidad para vincular las cepas del viñedo con los aislamientos detectados durante la vinificación y el envejecimiento. El número extremadamente bajo de Brettanomyces en las uvas, la distribución aleatoria de las cepas en los viñedos y las dificultades para aislarlas de sustratos ricos en azúcares han sido reportados previamente.
De manera similar, Garijo et al. realizaron un estudio durante tres cosechas consecutivas evaluando uvas y las etapas iniciales de fermentación, demostrando que la presencia de B. bruxellensis en las uvas es generalmente baja, lo que respalda la conclusión de que las uvas en sí no constituyen un punto crítico de infección por esta levadura. Por otro lado, Albertin et al. compararon cepas de B. bruxellensis detectadas en uvas y vino mediante análisis de microsatélites, mostrando que las cepas aisladas de uvas se agrupan junto con cepas provenientes del vino, lo que sugiere una fuerte conexión entre las uvas y la bodega.
Durante la fermentación alcohólica, no se detectaron poblaciones de Brettanomyces en ninguno de los vinos estudiados, a pesar de que las muestras fueron enriquecidas durante 60 días. Tras la finalización de la fermentación alcohólica, la bodega mantuvo los vinos entre 19 y 25 días a la espera de la fermentación maloláctica espontánea (FML). Durante este período, los vinos se mantuvieron en tanques a temperatura ambiente y sin la adición de SO₂, generando condiciones favorables para el incremento de las poblaciones de Brettanomyces.
Esto se evidenció en los recuentos positivos obtenidos en todos los vinos analizados en esta etapa. Estas poblaciones cultivables disminuyeron hasta niveles indetectables tras la sulfitación y antes del llenado en barricas. Resulta interesante destacar este período de espera para la FML como una etapa crítica para la bodega, ya que permite el crecimiento de Brettanomyces en un medio favorable y sin competencia de S. cerevisiae.
Además, al considerar los resultados de agrupamiento de biotipos, se observó que el clúster 3 incluyó aislamientos del final de la fermentación y de etapas previas al uso de barricas en tres de los vinos estudiados (Merlot, Malbec C y Malbec 2). Este agrupamiento podría sugerir, como se mencionó anteriormente, que la contaminación podría originarse en equipos compartidos de la bodega utilizados en etapas previas al envejecimiento, facilitando su diseminación.
Sin embargo, otros aislamientos de etapas previas al envejecimiento también se agruparon en clústeres que incluían aislamientos de un solo varietal. Por lo tanto, en esta etapa puede considerarse un origen mixto de las cepas presentes, algunas potencialmente provenientes del mosto de uva y otras introducidas mediante equipos contaminados de la bodega. La necesidad de considerar etapas tempranas de las operaciones en bodega como posibles fuentes de contaminación ha sido sugerida en estudios previos.
En el presente estudio, tras la finalización de la FML, los vinos fueron trasegados y prensados, y la mayoría recibió adición de sulfitos antes de ser transferidos a barricas, excepto el vino Merlot, previamente descrito. Este proceso dio lugar a recuentos negativos o muy bajos de células cultivables en los vinos durante los primeros meses de envejecimiento. Como se mencionó anteriormente, la adición de SO₂ controla la población de células cultivables y puede inducir el estado VBNC.
La mayoría de las muestras positivas para Brettanomyces durante el envejecimiento correspondieron al sexto mes. Esto es consistente con la práctica de la bodega de corrección de SO₂ (conocida como “rack and SO₂”), que implica reunir vinos del mismo lote almacenados en diferentes barricas en un tanque, ajustar los niveles de SO₂ y luego rellenar nuevamente las barricas. Además, tras esta práctica, los aislamientos de un mismo vino envejecido en diferentes barricas se agruparon dentro de los mismos clústeres, lo que sugiere una posible dispersión de la contaminación.
Por otro lado, los aislamientos de vinos positivos durante el envejecimiento se agruparon en diferentes clústeres, lo que sugiere la presencia de más de un biotipo en los vinos. Este hallazgo coincide con la literatura existente que destaca la diversidad de poblaciones de Brettanomyces durante el envejecimiento en barrica. La porosidad de las barricas, el vino residual y protocolos de limpieza inconsistentes pueden crear micronichos que permiten la persistencia y proliferación de ciertas cepas de B. bruxellensis.
Se ha demostrado que algunas cepas de B. bruxellensis pueden formar biopelículas en el vino, contribuyendo aún más a su persistencia. Un estudio sobre la dispersión geográfica y persistencia de aislamientos de Brettanomyces en bodegas reveló que ciertos genotipos pueden aislarse repetidamente en una misma bodega durante décadas, demostrando una capacidad de persistencia inesperada. La detección de múltiples clústeres en este estudio indica que más de una cepa se ha establecido con éxito en el ambiente de la bodega, lo que subraya la importancia de monitorear y controlar Brettanomyces en todas las etapas del proceso productivo.
Para evaluar si existía una contaminación generalizada dentro de la bodega, se incluyeron en el estudio cinco vinos que resultaron positivos para Brettanomyces durante el envejecimiento. Los aislamientos obtenidos de estos vinos se agruparon en dos clústeres relacionados, distintos de los aislamientos de los cuatro vinos originalmente estudiados. Este hallazgo sugiere que la bodega no alberga una cepa o grupo de cepas ampliamente diseminado que contamine de forma recurrente todos los vinos.
En cambio, distintos grupos de cepas parecen estar asociados con vinos específicos que envejecen en determinados conjuntos de barricas. Varios estudios han destacado el papel de los equipos de bodega, las barricas y los reservorios ambientales en la diseminación de Brettanomyces. Sin embargo, la ausencia de una cepa dominante en todos los vinos sugiere que los eventos de contaminación pueden ser localizados o estar vinculados a procesos específicos, como el trasiego del vino, el uso de barricas o diferencias en las prácticas de vinificación.
Conclusiones
Aunque el origen primario de Brettanomyces es la uva, habiendo ingresado inicialmente a la bodega por esta vía, los vinos contaminados con Brettanomyces no necesariamente tienen su fuente de contaminación en el viñedo. Otras fuentes de contaminación están más estrechamente relacionadas con las prácticas y el equipamiento de la bodega.
Este estudio demuestra que el equipamiento de la bodega puede actuar como vector para la introducción de cepas de Brettanomyces. Se identificaron dos etapas críticas: el uso compartido de equipos durante las etapas de fermentación, que facilita la diseminación de cepas de Brettanomyces entre distintos vinos, y la reutilización de barricas, que introduce nuevas cepas en los vinos durante el envejecimiento.
Este estudio resalta el impacto significativo de las prácticas de bodega en el crecimiento y la diseminación de Brettanomyces. Las prácticas que favorecen el crecimiento poblacional, como la realización de fermentación maloláctica espontánea (FML), generan períodos prolongados en los que los vinos se mantienen a temperatura ambiente sin adición de SO₂, proporcionando condiciones favorables para Brettanomyces.
Asimismo, las prácticas que favorecen la dispersión de la contaminación, como el procedimiento de «trasiego y sulfitado”, en el que los vinos de diversas barricas se combinan en un tanque, facilitan la diseminación de cepas desde barricas contaminadas hacia otras previamente no contaminadas.
Además, el movimiento del vino durante estas prácticas introduce oxígeno, lo que puede estimular el crecimiento de Brettanomyces. Si bien es imposible eliminar completamente Brettanomyces del ambiente de la bodega, su crecimiento puede prevenirse, y deben minimizarse aquellas prácticas que faciliten su desarrollo y diseminación.
Descargar estudio en: https://www.mdpi.com/2311-5637/11/4/175
Financiamiento: Esta investigación fue financiada por el Instituto Nacional de Tecnología Agropecuaria (INTA)—Convenio de Asistencia Técnica INTA-Bodega Comercial (nombre confidencial), código 24945, y por los proyectos “Instituto Nacional de Tecnología Agropecuaria, Números de Proyecto INTA-2023-PE-L01-I002 e INTA-2023-PE-L04-I119”. L.M.B. fue beneficiaria de una beca doctoral del Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET).
Contactos: sturm.maria@inta.gob.ar ; chimeno.valeria@inta.gob.ar ; mercado.laura@inta.gob.ar – magonzalez@fca.uncu.edu.ar – lerena.cecilia@inta.gob.ar; rojo.cecilia@inta.gob.ar; becerra.lucia@inta.gob.ar –combina.mariana@inta.gob.ar










