











Una reciente investigación de la Universidad de Extremadura analiza nuevas estrategias para potenciar el desempeño enológico de Torulaspora delbrueckii, una levadura no-Saccharomyces cada vez más utilizada por su aporte aromático y su capacidad para reducir la acidez volátil. El objetivo del trabajo fue mejorar su rendimiento fermentativo y aumentar el tamaño celular para favorecer su aplicación a escala industrial. A partir de técnicas de selección genética, hibridación y poliploidía transitoria, se lograron avances relevantes en dominancia fermentativa, viabilidad celular y estabilidad genética, consolidando el potencial de esta levadura como herramienta diferencial para la producción de vinos tranquilos y espumosos con perfiles sensoriales específicos.

Autores: Alberto Martínez, Luis Miguel Hernández y Manuel Ramírez, del Departamento de Ciencias Biomédicas (Área de Microbiología) de la Facultad de Ciencias de la Universidad de Extremadura; Felipe Molina, del Departamento de Bioquímica, Biología Molecular y Genética (Área de Genética) de la misma universidad; y María Luz Álvarez, de la Estación Enológica, adscrita a la Consejería de Medio Ambiente y Rural, Políticas Agrarias y Territorio de la Junta de Extremadura. Estudio publicado el 1/5/26 en la revista Enólogos “Incremento de la ploidía como estrategia para mejorar la eficacia fermentativa de Torulaspora delbrueckii».
El ciclo de vida de la mayoría de las levaduras no convencionales, como Torulaspora delbrueckii (Td), no está tan bien caracterizado como el de Saccharomyces cerevisiae (Sc). Generalmente se asume que Td es principalmente haploide, lo que se ha postulado como un factor que reduce su eficacia fermentativa en comparación con las estirpes diploides de Sc.
El análisis del ciclo de vida de varias estirpes vínicas Td indica que son predominantemente diploides durante la fase de crecimiento exponencial en medio rico. Sin embargo, la mayoría de las células se vuelven haploides al dejar de crecer y entrar en la fase estacionaria. Cuando se transfieren a medios deficientes en nutrientes, estas células haploides se vuelven pleomórficas, aumentan de tamaño y pasan a estados diploides o poliploides. Este aumento de ploidía, que se deriva primariamente de mitosis supernumerarias sin la posterior citocinesis, es seguido por la esporulación.
Se observó una respuesta similar en levaduras que se mantienen viables durante la segunda fermentación del vino base para la elaboración de cava, así como durante el crecimiento en mosto sintético suplementado con etanol. Es crucial destacar que esta respuesta no se observó en levaduras Sc en ninguna de las condiciones experimentales mencionadas, lo que sugiere que se trata de una adaptación específica de Td a las condiciones estresantes de fermentación.
Esta adaptabilidad podría ser el mecanismo que permite a las levaduras Td permanecer viables y metabólicamente activas durante períodos más largos en la fermentación vínica. En consecuencia, en este estudio se han diseñado y evaluado procedimientos destinados a inducir el aumento del tamaño celular y la ploidía de estirpes haploides Td. Los inóculos Td con ploidía aumentada mostraron una mejora significativa en la eficacia fermentativa en comparación con los inóculos haploides de las mismas estirpes.
Introducción
La levadura Torulaspora delbrueckii (Td) está despertando un creciente interés como cultivo iniciador en la elaboración de vinos, debido a su demostrada capacidad para mejorar la complejidad del perfil aromático del vino y reducir la acidez volátil. Sin embargo, su menor rendimiento fermentativo y tamaño celular en comparación con Sc condicionan su uso a escala industrial. Para superar estas restricciones, se han implementado diversos enfoques de mejora genética, que incluyen la selección de mutantes espontáneos resistentes a condiciones de estrés y la obtención de estirpes panaderas diploides. Así se ha logrado incrementar tanto su eficacia fermentativa como su viabilidad.
Asimismo, se han explorado estrategias de hibridación interespecífica entre Td y Sc, obteniéndose clones con propiedades enológicas mejoradas y suficiente estabilidad genética para aplicaciones comerciales. La investigación reciente sobre la ploidía y el ciclo de vida de Td revela comportamientos reproductivos complejos y dinámicos en respuesta a condiciones de estrés, como las encontradas durante la fermentación.
Estos mecanismos biológicos pueden ser aprovechados estratégicamente para optimizar las aplicaciones enológicas de la levadura. Todos estos avances amplían significativamente las posibilidades de uso de Td en la industria vitivinícola, ofreciendo un potencial diferenciador para el desarrollo de vinos con características sensoriales únicas y controladas.
Materiales y métodos
Se emplearon diversas estirpes de Td y Sc, incluyendo estirpes comerciales de referencia, mutantes espontáneos seleccionados y clones hibridados interespecíficos. Las estirpes Td, caracterizadas por su baja capacidad para fermentar maltosa, fueron analizadas para su resistencia a estrés (SO2, etanol, alta presión de CO2), actividad killer, producción de H2S y capacidad de esporulación.
Para el cultivo y la propagación, se emplearon medios estándares como YEPD y Extracto de Malta. Para inducir la esporulación se utilizó medio específico SPO. Los ensayos de fermentación se realizaron en mosto sintético (220 g/L azúcares reductores totales, 200 mg/L de nitrógeno asimilable) y en vino base sintético con 8–9 % (v/v) de etanol, ajustado a pH 3.1.
La cinética de fermentación se monitorizó midiendo periódicamente los grados Brix. Asimismo, se determinaron la viabilidad celular (método azul de metileno) y la proporción de las levaduras inoculadas mediante marcadores genéticos, como resistencia a cicloheximida (cyhR) y el análisis de restricción de ADN mitocondrial.
El análisis de la distribución de los distintos tipos de células durante el ciclo celular y el nivel de ploidía se realizó mediante citometría de flujo y microscopía confocal, previa tinción nuclear. Para confirmar la identidad genética de las distintas estirpes se secuenciaron las regiones ITS del ADN ribosomal. La combinación de estas herramientas moleculares con la caracterización fenotípica y fermentativa permitió la evaluación integral del potencial enológico y biotecnológico de las estirpes de Td estudiadas.
Resultados y discusión
Ploidía de Td durante el crecimiento vegetativo y la esporulación
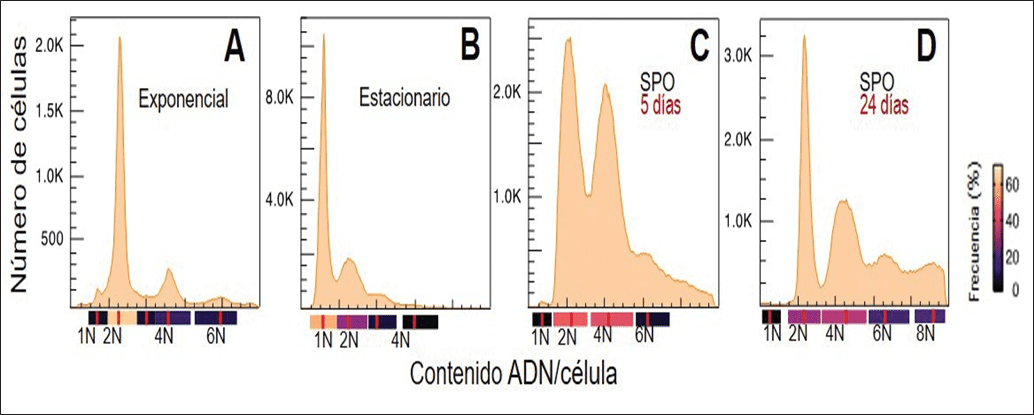
El análisis de citometría de flujo reveló una dinámica de la ploidía dependiente de la fase de crecimiento en (Td). Durante la fase de crecimiento exponencial en medio YEPD, la mayoría de las células de la estirpe de referencia Td EX1180 presentaron un estado diploide (2N), con una fracción menor de células tetraploides (4N) y hexaploides (6N), mientras que las poblaciones haploides (1N) fueron prácticamente indetectables.
Al alcanzar la fase estacionaria, la mayoría de las células transitaron al estado haploide (1N), aunque se mantuvieron algunas células diploides (2N) y triploides (3N) residuales. Este comportamiento de transición (2N a 1N) se repitió en las 19 estirpes analizadas, siendo el estado haploide final comparable al observado en estirpes haploides de Sc (Figuras 1A y B).
Al transferir las células de Td a medio de esporulación, las estirpes silvestres mostraron un incremento progresivo en la ploidía, con desaparición casi total de células haploides y aparición de células hexaploides (6N) y octaploides (8N) (Figuras 1C y D). Paralelamente, se identificaron células de gran tamaño con morfologías inusuales tales como tubos de conjugación y estructuras en forma de coronas de yemas (“Kronenbildung”) (Figuras 2B, C y G), ausentes en estirpes diploides de Sc.
La estrecha relación entre la ploidía aumentada y el ciclo reproductivo se confirmó con la estirpe deficiente en esporulación, Td SpoDef9, la cual no mostró aumento en la ploidía. Este resultado sugiere una relación directa entre el potencial de esporulación y la capacidad de transición a estados de ploidía aumentada como respuesta al estrés nutricional.
Los resultados descritos demuestran que ciertas condiciones de estrés (como la limitación de nutrientes, el etanol o el frío) inducen transiciones estables a estados diploides o poliploides, revelando una flexibilidad de ploidía clave para su adaptación.
Efecto del estrés fermentativo y de las condiciones de almacenamiento sobre la ploidía deTd
Durante la segunda fermentación de vino base sintético (simulando la elaboración de cava y espumosos), las poblaciones de Td mostraron un aumento notable en la frecuencia de células pleomórficas de gran tamaño. Este patrón morfológico se reprodujo en condiciones simuladas de estrés fermentativo (mosto sintético con etanol al 5 %). El contenido de ADN de estas células se confirmó cuantitativamente mediante citometría de flujo, estableciendo una correlación entre el estrés alcohólico y la poliploidía inducida.
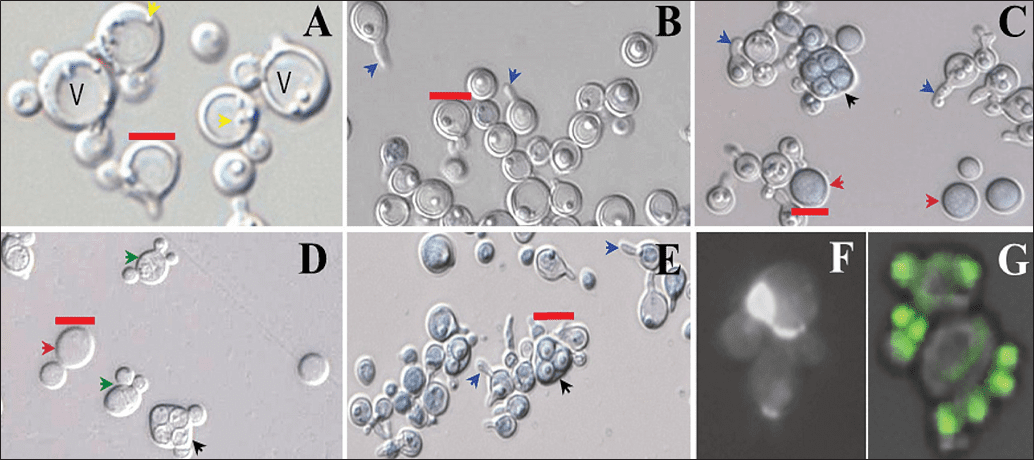
Se evaluaron diversas condiciones de cultivo y almacena-miento con el fin de generar inóculos de Td con mayor tamaño celular y ploidía estable buscando un protocolo industrialmente viable. Entre las respuestas celulares se incluyen: autofagia, evidenciada por la presencia de autofagosomas y vacuolas de gran tamaño (Figura 2A); acumulación de lípidos, formación de proyecciones de apareamiento y esporulación incipiente (Figura 2B), observación de aparentes tubos conjugativos y esporulación final (Figuras 2C, D y E), y ploidía inducida con la presencia de células con mitosis supernumerarias sin citocinesis (Figuras 2F y G).
Las condiciones que resultaron más efectivas para inducir células con ploidía aumentada fueron: inóculos de Td cultivados durante 4 días en caldo YEPD a 30º C, seguidos de transferencia a caldo SPO o extracto de malta al 1,7 % y almacenamiento a 15 °C (Td-SPO o Td-Malta respectivamente).
Efecto de los inóculos de Td con ploidía aumentada sobre la fermentación de mosto y vino base sintético
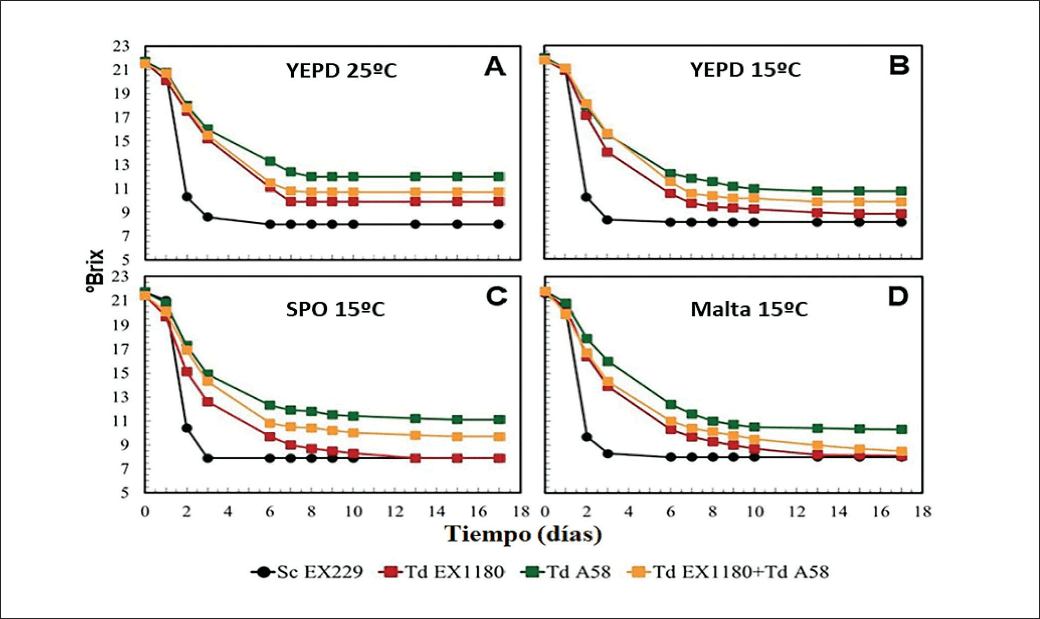
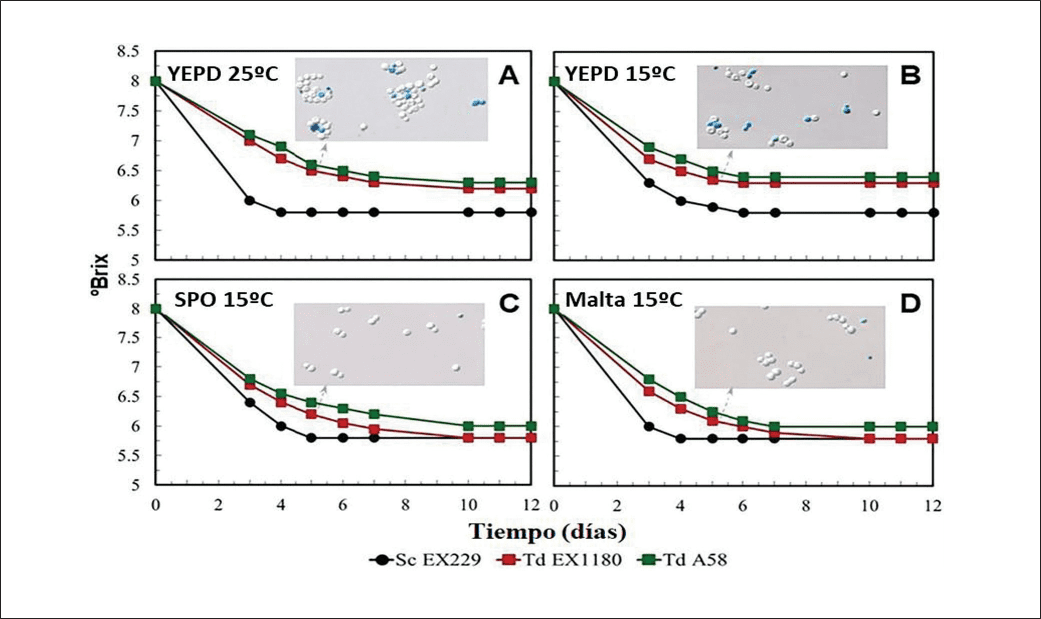
En mosto sintético las poblaciones de Td-SPO y Td-Malta dominaron todas las fermentaciones y mejoraron significativamente la cinética fermentativa en comparación con los controles (cultivos almacenados en YEPD) (Figuras 3A y B). La eficacia fermentativa en mosto sintético de las levaduras Td fue notablemente superior cuando el tratamiento del inóculo fue de cuatro días a 15 °C (Figura 3B) comparado con el tratamiento a 25º C (Figura 3A). En la estirpe de referencia Td EX1180,el rendimiento fermentativo de los inóculos Td-SPO (Figura 3C) y Td-Malta (Figura 3D) fue capaz de igualar al de la estirpe control Sc. Este resultado destaca la capacidad de la poliploidía inducida para superar una de las principales limitaciones de Td, fermentación deficiente respecto a Sc.
En las condiciones más exigentes del vino base sintético (8-9% etanol), ningún inóculo de Td fue tan eficiente como la levadura control Sc, y no hubo mejora de la eficacia fermentativa de Td con los inóculos Td-YEPD a 15 °C (Figura 4B) frente a 25 °C (Figura 4A). Sin embargo, en comparación con sus propios controles, los inóculos Td-SPO (Figura 4C) y Td-Malta (Figura 4D) superaron consistentemente la eficacia fermentativa de los inóculos del control Td-YEPD.
La viabilidad celular se identificó como un factor clave, ya que el porcentaje de células muertas fue significativamente menor en los inóculos Td-SPO y Td-Malta en comparación con los inóculos del control Td-YEPD. Esta correlación directa entre viabilidad y eficiencia fermentativa es crítica, demostrando que los inóculos de ploidía aumentada mantienen una mejor integridad celular bajo estrés y tendieron a completar la fermentación en plazos aceptables.
La implementación de este protocolo podría optimizar procesos como la segunda fermentación de vinos espumosos o la fermentación de mostos de alto grado, reduciendo la necesidad de etapas de adaptación previa o coinoculación con Sc.
Conclusiones
Las poblaciones de T. delbrueckii exhiben una notable flexibilidad de ploidía. Si bien predominan las células haploides (1N) en fase estacionaria, el estrés fermentativo induce la formación de células pleomórficas con un aumento significativo en tamaño y ploidía. Este mecanismo se confirma como una estrategia adaptativa biológica clave para mejorar la resistencia y la supervivencia de Td en el entorno enológico.
El uso de inóculos de Td con ploidía aumentada, generados bajo condiciones controladas (Td-SPO y Td-Malta), mejoró significativamente la cinética fermentativa tanto en mosto sintético como en vino base sintético. En mosto, esta mejora permitió a la estirpe Td EX1180 igualar el rendimiento de las estirpes de referencia de Sc.
Estos hallazgos demuestran que la inducción transitoria de poliploidía en Td es una estrategia biotecnológica robusta para la mejora de inóculos de estirpes vínicas. No solo favorece la dominancia y eficiencia de Td en fermentaciones de vinos tranquilos y espumosos, sino que el aumento del tamaño celular asociado, podría ofrecer beneficios adicionales, como la optimización de los procesos de filtrado y deshidratación del inóculo a escala industrial.
rabajo financiado por las subvenciones GR21062, IB16132 e IB20069 de la Junta de Extremadura, AGL2017-87635-R de MICIU/AEI/10.13039/501100011033 (cofinanciada por FEDER Una forma de hacer Europa) y GR24105 de la Junta de Extremadura (85 % de la Unión Europea, Fondo de Desarrollo Regional y Junta de Extremadura. Ministerio de Hacienda).








